โปรตีนเป็นสารชีวโมเลกุลขนาดใหญ่ซึ่งเป็นส่วนประกอบโครงสร้างพื้นฐานของสิ่งมีชีวิตทุกชนิด พวกมันคือสารประกอบโมเลกุลขนาดใหญ่ที่มีกรด α-อะมิโนตกค้างและพันธะเอไมด์ หรือที่เรียกว่าพันธะเปปไทด์ น้ำหนักโมเลกุลของโปรตีนเกิน 10,000 ดาลตัน (ดา) เปปไทด์ที่มีน้ำหนักโมเลกุลต่ำกว่าประกอบด้วยกลุ่มที่แยกจากกันซึ่งเรียกว่าโอลิโกเปปไทด์ โปรตีนทั่วไปประกอบด้วยกรดอะมิโนมากถึง 20 ชนิด ซึ่งทั้งหมด (ยกเว้นไกลซีน) มีสเตอริโอเคมีคล้ายกับกรดแอล-น้ำตาล โมเลกุลเคมีเหล่านี้สามารถจำแนกได้เป็นสองกลุ่มทั้งนี้ขึ้นอยู่กับองค์ประกอบ อันดับแรกรวมถึงโปรตีนอย่างง่าย เช่น โปรตีนที่ไฮโดรไลซ์เป็นกรดอะมิโนเท่านั้น กลุ่มนี้รวมถึงเช่น เซรั่มอัลบูมิน กลุ่มที่สองประกอบด้วยโปรตีนเชิงซ้อนที่ไฮโดรไลซ์เป็นผลิตภัณฑ์ที่มีกรดอะมิโนและสารประกอบอื่นๆ เช่น คาร์โบไฮเดรต ไขมัน และกรดนิวคลีอิก
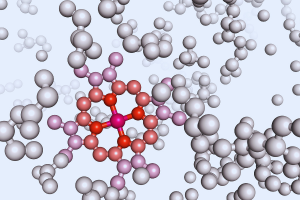
องค์ประกอบทางเคมีของโปรตีน
พารามิเตอร์ทางเคมีพื้นฐานที่กำหนดกลุ่มของสารประกอบที่กำหนดคือองค์ประกอบทางเคมีของพวกมัน จากการวิเคราะห์องค์ประกอบพบว่าโครงสร้างของแต่ละตัวแทนของกลุ่มโปรตีนประกอบด้วยคาร์บอน (50-55%) ไฮโดรเจน (6-7%) ออกซิเจน (20-23%) ไนโตรเจน (12-19 %) และฟอสฟอรัสในปริมาณที่น้อยกว่า (0-6%) และกำมะถัน (0.2-3%)
การจำแนกประเภทของโปรตีนเนื่องจากรูปแบบสามมิติ
โปรตีนยังจัดประเภทเป็น ไฟบริลลาและโกลบูลาร์ ซึ่งอ้างอิงโดยตรงกับรูปแบบสามมิติของพวกมัน โปรตีนไฟบริลลาร์สร้างจากสายโซ่โพลีเปปไทด์ที่สร้างเส้นใยยาว พวกมันทนทานและไม่ละลายในน้ำ ดังนั้นพวกมันจึงทำหน้าที่เป็นวัสดุก่อสร้างสำหรับเนื้อเยื่อโครงสร้าง เช่น เส้นเอ็น กีบเท้า เล็บ เนื้อเยื่อเกี่ยวพันของหลอดเลือดและกล้ามเนื้อ ตัวแทนที่พบบ่อยที่สุดของกลุ่มนี้ ได้แก่ คอลลาเจนและครีเอทีน รวมถึงไฟบริโนเจน อีลาสติน และไมโอซิน โปรตีนทรงกลมหรือโกลเมอรูลเป็นโมเลกุลในรูปของคอมแพคพับคล้ายทรงกลม โปรตีนเหล่านี้มักจะละลายได้ดีในน้ำและเคลื่อนย้ายได้อย่างอิสระภายในเซลล์ โครงสร้างประเภทนี้เป็นเรื่องปกติสำหรับเอนไซม์ส่วนใหญ่ที่รู้จัก เฮโมโกลบิน อิมมูโนโกลบูลิน อินซูลิน และไรโบนิวคลีเอส การใช้โปรตีนทรงกลมรวมถึงกระบวนการขนส่งออกซิเจน ปฏิกิริยาภูมิคุ้มกัน และกฎระเบียบของฮอร์โมนและเอนไซม์ รวมถึงเมแทบอลิซึมของกลูโคสและการสังเคราะห์ RNA
คุณสมบัติแอมโฟเทอริกของโปรตีน
กรดอะมิโนที่แตกตัวเป็นไอออนในปริมาณค่อนข้างมากที่มีอยู่ในโปรตีนทรงกลมทำให้พวกมันมีคุณสมบัติ ในการแก้ปัญหาทั้งกรดและด่าง คุณสมบัติของพวกเขาขึ้นอยู่กับสภาพแวดล้อม ในสารละลายที่เป็นกรด เนื่องจากมีไฮโดรเจนไอออนจำนวนมาก การแตกตัวของกลุ่มที่เป็นกรดจะย้อนกลับ ทำให้โมเลกุลของโปรตีนกลายเป็นไอออนบวก ในสถานการณ์ตรงกันข้าม โมเลกุลเป็นไอออน – ในสภาพแวดล้อมที่เป็นด่าง กลุ่มอัลคาไลน์จะสูญเสียประจุไฟฟ้า เนื่องจากลักษณะสองอย่างนี้ โปรตีนสามารถแยกตัวออกได้สองวิธี – ในทางที่เป็นกรดและด่าง ระดับการแยกตัวและจำนวนของประจุจะขึ้นอยู่กับค่า pH และชนิดของ กรดอะมิโน ที่มีอยู่ในโครงสร้างโดยตรง นอกจากนี้ยังมี จุดไอโซอิเล็กทริก เช่น ค่า pH ของสิ่งแวดล้อมซึ่งมีประจุบวกและลบเท่ากัน และโมเลกุลของโปรตีนจะกลายเป็นสวิตเตอร์ไอออน ค่านี้เป็นลักษณะเฉพาะของโปรตีนแต่ละชนิดและทำให้สามารถแยกออกได้ ที่จุดไอโซอิเล็กทริก:
- สังเกตค่าที่น้อยที่สุดของประจุไฟฟ้าและค่าการนำไฟฟ้าต่ำสุด
- โปรตีนแสดงการเคลื่อนที่ต่ำสุด ดังนั้นโปรตีนส่วนใหญ่จึงตกตะกอนหรือกลายเป็นโซล/เจล
- คุณสมบัติต่างๆ เช่น ความหนืด ความสามารถในการพองตัว ความสามารถในการละลาย แรงดันออสโมติกต่ำที่สุด
โครงสร้างของโปรตีน
กลุ่มสารประกอบนี้มีลักษณะเป็นโครงสร้างขนาดใหญ่มากในสี่ระดับที่แตกต่างกัน พูดง่ายๆ ก็คือ โปรตีนคือลำดับของกรดอะมิโนที่จับกัน นี่คือระดับพื้นฐานที่สุดของการจัดเรียง – โครงสร้างหลัก โครงสร้างทุติยภูมิ เป็นการจัดเรียงตัวปกติ ซึ่งเป็นผลมาจากการโค้งงอของแกนโพลีเปปไทด์ และจากโครงสร้างสามมิติของมัน นอกจากนี้ โครงสร้างตติยภูมิ เกี่ยวข้องกับการจำแนกประเภทตามการพับของโมเลกุลเป็นรูปร่างสุดท้าย โครงสร้างควอเทอร์นารี อธิบายการก่อตัวของมวลรวมขนาดใหญ่ขึ้นโดยโมเลกุลโปรตีนที่กำหนด
เอนไซม์
เอนไซม์เป็น กลุ่มของโปรตีนขนาดใหญ่ที่ทำหน้าที่เป็นตัวเร่งปฏิกิริยาในปฏิกิริยาทางชีวภาพหลายชนิด พวกมันแตกต่างจากตัวเร่งปฏิกิริยาเคมีที่ใช้ในห้องปฏิบัติการเนื่องจากการกระทำเฉพาะของมัน โดยปกติแล้ว เอนไซม์หนึ่งตัวสามารถเร่งปฏิกิริยาได้เพียงหนึ่งปฏิกิริยาของสารประกอบเพียงชนิดเดียวเท่านั้น ซึ่งเรียกว่าสารตั้งต้น ตัวอย่างเช่น ระบบทางเดินอาหารของมนุษย์มีอะไมเลสซึ่งเร่งปฏิกิริยาไฮโดรไลซิสของแป้งให้เป็นกลูโคสเท่านั้น และไม่เร่งปฏิกิริยาไฮโดรไลซิสของเซลลูโลสหรือพอลิแซ็กคาไรด์อื่นๆ นอกจากนี้ยังมีเอนไซม์อื่นๆ เช่น ปาเปน ซึ่งทำหน้าที่กับสารตั้งต้นทั้งกลุ่ม ในกรณีนี้จะเร่งปฏิกิริยาไฮโดรไลซิสของพันธะเปปไทด์หลายชนิด เช่นเดียวกับตัวเร่งปฏิกิริยาทางเคมี เอนไซม์จะไม่รบกวนสมดุลคงที่ของปฏิกิริยา แต่จะลดพลังงานกระตุ้นลงเท่านั้น ซึ่งส่งผลให้เกิดการเร่งความเร็วของกระบวนการ
การสูญเสียโปรตีน
โปรตีนทรงกลมมีโครงสร้างระดับตติยภูมิที่สมดุลโดยปฏิกิริยาภายในโมเลกุลที่อ่อนแอ มันง่ายมากที่จะทำลายมัน เช่น การเปลี่ยนแปลงเล็กน้อยของอุณหภูมิหรือค่า pH ซึ่งส่งผลให้โปรตีนเสียสภาพไปในที่สุด อย่างไรก็ตาม เงื่อนไขเหล่านี้มีการบุกรุกน้อยที่สุดจนพันธะโควาเลนต์ไม่แตก ด้วยเหตุนี้ โครงสร้างโพลีเปปไทด์หลักจึงไม่เปลี่ยนแปลง อย่างไรก็ตาม โครงสร้างแต่ละส่วนไม่เหมือนกัน ซึ่งนำไปสู่การตีแผ่จากรูปทรงกลมและเปลี่ยนให้เป็นมัดที่ยุ่งเหยิง อย่างไรก็ตาม ยังมีปัจจัยอื่นๆ ที่ทำให้โปรตีนเสียสภาพธรรมชาติ เช่น รังสียูวี การสั่นอย่างรุนแรง ความดันสูง และปัจจัยทางเคมีหลายอย่าง รวมถึงกรดแก่และเกลือของโลหะหนัก การเสียสภาพธรรมชาติของโปรตีนทำให้เกิดการเปลี่ยนแปลงอย่างมากใน คุณสมบัติทางกายภาพและทางเคมี ของโปรตีน ความสามารถในการละลายลดลงอย่างมาก ซึ่งสามารถสังเกตได้ เช่น เมื่อปรุงอาหารไข่: อัลบูมินจะคลี่ออกและจับตัวเป็นก้อนเป็นก้อนสีขาวที่ไม่ละลายน้ำในรูปของโปรตีนนมเปรี้ยว เนื่องจากการเสียสภาพธรรมชาติ เอ็นไซม์ส่วนใหญ่สูญเสียกิจกรรมทางชีวภาพ เนื่องจากโครงสร้างตติยภูมิที่สำคัญถูกทำลาย หลังการเสียสภาพธรรมชาติ กิจกรรมของกลุ่มสารเคมีที่สัมผัสเพิ่มขึ้น มุมการหมุนของระนาบของแสงโพลาไรซ์เพิ่มขึ้น และความไวต่อเอ็นไซม์สลายโปรตีนเพิ่มขึ้น โดยปกติแล้ว กระบวนการเปลี่ยนสภาพธรรมชาตินั้นไม่สามารถย้อนกลับได้ แต่การเสียสภาพธรรมชาติที่เกิดขึ้นเองนั้นเกิดขึ้นในระยะเริ่มต้นของการเผยตัวของโมเลกุล จากนั้นเอนไซม์ก็จะคืนกิจกรรมทางชีวภาพที่สูญเสียไปก่อนหน้านี้ จากการสังเกตนี้สรุปได้ว่าโครงสร้างระดับตติยภูมิของพวกมันกลับคืนสู่รูปแบบที่มั่นคงอย่างสมบูรณ์

